Introduction
In the past 20 years great stress has been laid on the role of oxidative stress caused by reactive oxygen species (ROS), differences in antioxidant mechanisms among individuals, especially in age-dependent diseases such as cancer, arthritis, arteriosclerosis, neurodegenerative disorders and others (1,2).
Antioxidant defense is important in the removal of free radicals, providing the maximal protection of biological sites such as thiol groups which are part of active sites in some metabolizing enzymes (2,3). A good antioxidant should specifically quench free radicals, chelate redox metals, interact with other antioxidants within an antioxidant network, have a positive effect on gene expression, be readily absorbed, have a concentration in tissues and biofluids at a physiologically relevant level, act in both aqueous and/or membrane domains (2). The most efficient enzymatic antioxidants involve superoxide dismutase, catalase and glutathione peroxidase. Non-enzymatic antioxidants involve thiol antioxidants (glutathione, thioredoxin and lipoic acid), vitamin C, vitamin E, carotenoids, natural flavonoids, melatonin and other compounds (selenium) (4).
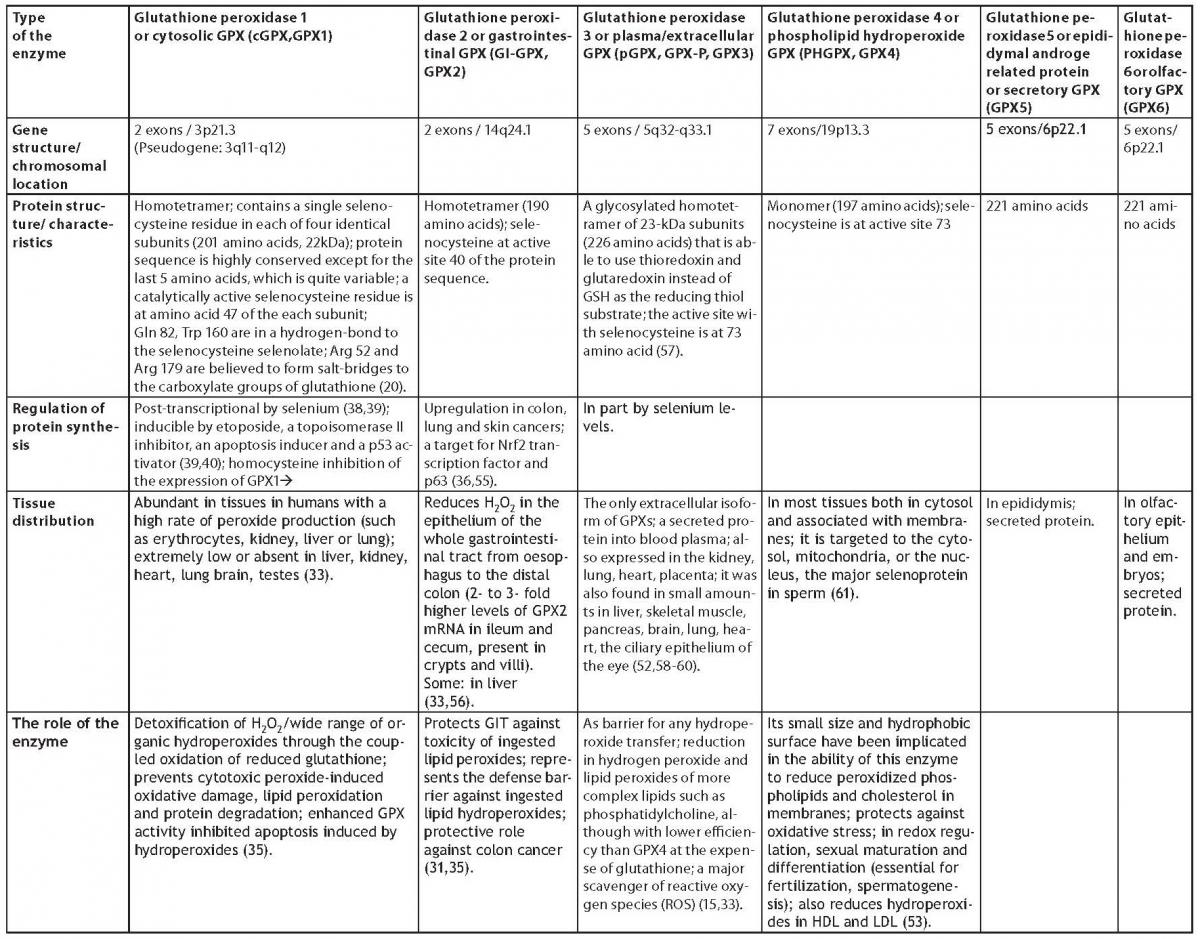
In this review the focus is placed on a group of enzymes, glutathione peroxidases (GPXs), which are the major enzymes in the antioxidative defense mechanism depending on glutathione. The specific biochemical properties, synthesis and the role of each GPX isoenzyme will be described in biological processes. At least 7 types of GPXs have been identified (5,6), and their properties will be collected in a table divided into the following types: cytosolic- (cGPX or GPX1), gastrointestinal- (GI-GPX or GPX2), plasma- (pGPX or GPX3), phospholipid hydroperoxide- (PHGPX or GPX4) glutathione peroxidase, GPX5 and GPX6. Furthermore, non-enzymatic antioxidants, included in the activity of GPXs, will be reviewed.
Biosynthesis of glutathione peroxidases
Biosynthesis of glutathione peroxidases is similar to biosynthesis of all selenoproteins which depends on the availability of selenium (Se). It was established in 1973 that Se is a structural component of the active center of the animal enzyme cellular glutathione peroxidase (GPX1) (6,7). Since then, 30 new selenoproteins have been identified, 15 of them were purified and their biological function was characterized (7-9).
Selenium is incorporated as selenocysteine (Sec) into the active site of a growing polypeptide chain encoded by UGA. This cotranslational incorporation of Sec into selenoproteins presents significant problems to the cell which must recognize the UGA as a Sec codon rather than a STOP translation signal (10,11,12).
The cloning of GPX1 has led to the identification of a specific eukaryotic Sec insertion sequence (SECIS) element as a stem-loop structure located in the 3ž untranslated regions (UTR) of the glutathione peroxidase mRNAs. The SECIS element, which is in the active site, is the signal that recodes the in-frame UGA from a STOP to a Sec codon (13). Evidently, this complicated sequence provides ample opportunity for a posttranscriptional regulation of selenoprotein biosynthesis by selenium. However, selenium deficiency results in premature chain termination at the UGA codon as well as in increased degradation of the selenoprotein mRNAs (14).
The minimal region required for SECIS function are conserved 5’ A/GUGA and 3’ GA sequences. This region has been subsequently proven to form non-Watson-Crick base pairs: purine pairs between the GA at the 5’ base of the stem (in the conserved A/GUGA sequence) and the GA at the 3’ base, and pyrimidine pairs flanking these two. Similar non-standard base-pairing features turned out to serve as binding sites for several sequence- and structure- specific RNA-binding proteins. In SECIS elements, the conserved adenosines are contained in a simple open loop (10,15) (Figure 1).
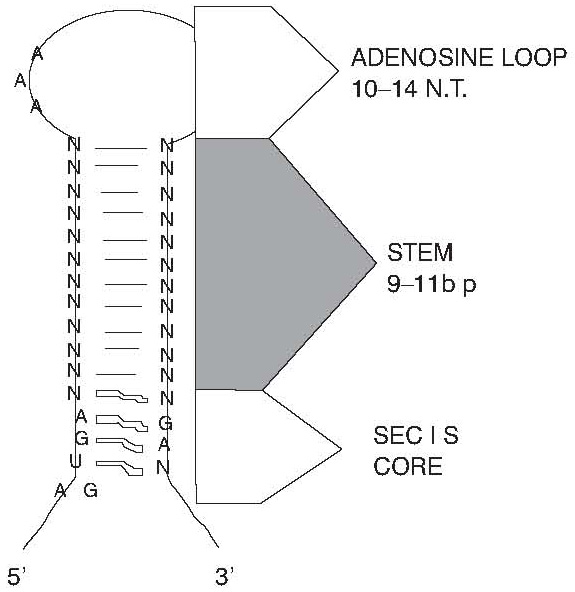
Figure 1. This figure shows the form of I SECIS element (GPX1). Conserved sequence and structural features include the SECIS core nucleotides (A/GUGA and GA), the stem length, and conserved adenosines in a terminal loop (marked as adenosine loop). The thin lines indicate Watson-Crick base pairs while the bold lines designate non-Watson-Crick pairing. The Form II SECIS element is the one with two stems, separated by a polyA region and is typical for GPX3 and GPX4.
The Sec insertion in eukaryotes requires dedicated translation factors including a Sec tRNA and elongation factor in addition to the RNA element in the 3’-untranslated portion of the mRNA that directs Sec incorporation in response to all in-frame UGA codons. In mammalian cells, this process is highly regulated and responsive to selenium availability, both at the levels of RNA stability and translation (11,13,14). Crucial for the Sec incorporation is a SECIS-specific binding protein, termed SECIS binding protein 2 (SBP2) which also binds to the elongation factor EFsec with specificity for selenocysteyl-tRNA (tRNA(Ser)Sec). SBP2 binds to a conserved, non-Watson-Crick base-paired region in the stem of the SECIS element and remains bound through multiple cycles of selenoprotein translation. tRNA(Ser)Sec is aminoacylated with serine which is subsequently converted to Sec (13,16,17).
As shown in Figure 2, the SECIS element recruits the SBP2 in the nucleus and the SECIS-SBP2 complex could recruit the EFsec-tRNA complex and deliver it to a ribosome in the coding region. Because the SECIS element is located in the 3’ UTR in eukaryotes, not in the coding region as in prokaryotes, it obviates the need for dissociation and reassociation of the SECIS-SBP2 complex with each incorporation cycle. This scheme could allow a rapid reformation of SECIS-SBP-EFsec-tRNA complexes from two individual RNA-protein complexes after each EFsec-tRNA delivery cycle reloading for the next approaching ribosome. This would also be advantageous in the translation of a protein containing multiple Sec residues such as selenoprotein P (5,11,13).
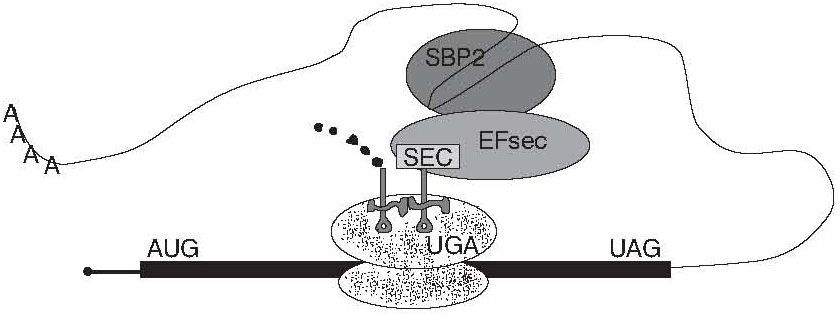
Figure 2. Sec incorporation of GPX directed from 3’ UTR. The open reading frame of an eukaryotic selenoprotein mRNA is depicted by the black bar with a ribosome (dots) decoding the UGA Sec codon. The UTRs are indicated by the thin black line. The SECIS-SBP2-EFsectRNA complex is shown assembled in the 3-UTR and looping back to the ribosome.
Table 1. Genes required for selenoprotein synthesis in Eukaryotes

A specific t-RNA(ser)sec is first loaded with serine, then transformed into selenocysteyl-tRNA(ser)sec with selenophosphate as the selenium donor and bound with its anticodon region to a UGA codon of the mRNA (14,15).
Interestingly, the stability of the mRNAs appears to correlate with the efficiencies of the pertinent SECIS elements in stop-codon suppression or selenocysteine incorporation. This observation has led to an intriguing hypothesis that SelC binding to the SECIS motif has dual effect in eukaryotes, resulting in the recoding of the UGA codon and stabilization of the mRNA in a selenium-dependent manner (18).
The structure of GPXs
Although GPX1, GPX3 and GI-GPX2 are homotetramers, the GPX4 is a monomer with a molecular size smaller than the subunits of other glutathione peroxidases. Due to their small size and hydrophobic surface, GPX4s have the ability to react with complex lipids in membranes (19).
The structure of mammalian GPXs was also analyzed by computer-assisted molecular modelling. The obtained models show that essential steps of catalysis are three distinct redox changes of the active site selenium, which in the ground state presents itself at the surface of selenoperoxidases as the center of a characteristic triad built by selenocysteine, glutamine and tryptophan. In GPX, four arginine residues and a lysine residue provide an electrostatic architecture which in each reductive step directs the donor substrate glutathione (GSH) towards the catalytic center in such a way that its sulfhydryl group must react with the selenium moiety. Moreover, cosubstrate binding mechanisms are unique for the classical type of GPX1 but cannot operate in GPX3 and GPX4 (11) (Figure 3).
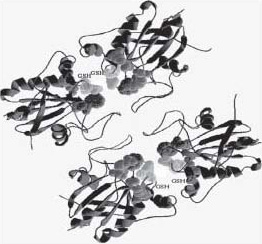
Figure 3. Structural model of GPX1 as a homotetramer (spheres with vertical lines indicate the active sites - selenocysteine at amino acid 47, the brightest grey spheres: Gln 82, white dotted spheres: Trp 160, spheres with horizontal lines: Arg 52 and Arg 179).
Analysis of selenoproteome characterized the functions and sequence of 6 glutathione peroxidases (GPXs) in mammals: cytosolic GPX (cGPX, GPX1), the first identified mammalian selenoprotein (6,15), phospholipid hydroperoxide GPX (PHGPX, GPX4) which was first described in 1982 and later verified as selenoprotein by sequencing (21,22), and other sequenced plasma GPX (pGPX, GPX3)(23), gastrointestinal GPX (GI-GPX, GPX2)(24) and, in humans, GPX6 which is restricted to the olfactory system (19).
Glutathione and selenium are important in GPX-related antioxidative defense mechanisms
Glutathione peroxidase is involved in protection against oxidative stress, and thus uses glutathione as a substrate. Glutathione also acts as a substrate in other detoxifying enzymes against oxidative stress, such as glutathione transferases. It participates in amino acid transport through the plasma membrane, scavenges hydroxyl radical and singlet oxygen directly, detoxifying hydrogen peroxide and lipid peroxides by the catalytic action of GPX. Glutathione is able to regenerate the most important antioxidants, vitamins C and E back to their active forms (2).
Due to cysteine that comprises a thiol group, glutathione is an important intracellular non-enzymatic antioxidant. Glutathione is highly abundant in the cytosol (1-11mM), nuclei (3-15mM), and mitochondria (5-11mM) and is a major soluble antioxidant in cell compartments (2,25).
The intracellular content of glutathione depends on environmental factors and functions as a balance between its utilization and synthesis. Exposure to ROS (involving H2O2)/RNS, or to compounds which can generate ROS, can increase the content of GSH by increasing the rate of GSH synthesis (2).
Significantly, GPX competes with catalase for H2O2 as a substrate. Glutathione redox cycle is a major source of protection against mild oxidative stress, whereas catalase becomes increasingly important in protection against severe oxidative stress (26).
However, in animal cells, and especially in human erythrocytes, the principal antioxidant enzyme for H2O2 detoxification has for a long time been considered to be GPX, as catalase has much lower affinity for H2O2 than GPX (2,27).
A large number of studies have established an association between cancer incidence and various disorders of GSH-related enzyme functions, while glutathione S-transferases (GSTs) have been more frequently reported with regard to alterations of glutathione peroxidases (2,3,25).
Selenium, as part of the active site in GPXs, is an essential micronutrient shown to reduce colon cancer incidence and preneoplastic aberrant crypt foci in animal models (27, 28, 29). It has also been implicated in possible chemoprevention of some cancers. In human studies, data have indicated that selenium levels are inversely associated with cancer mortality and incidence (20,30,31).
Thus, it appears to function as an antimutagenic agent, preventing the malignant transformation of normal cells. These protective effects of Se seem to be primarily associated with the activity of glutathione peroxidases. GPX1 levels are particularly responsive to fluctuations in selenium levels compared with other selenoproteins (6).
Specific biological properties of human glutathione peroxidases
1. Functions of glutathione peroxidases
Although their expression is ubiquitous, the levels of each isoform vary depending on tissue type. All glutathione peroxidases reduce hydrogen peroxide and alkyl hydroperoxides at the expense of glutathione. Their specificities for hydroperoxide substrate, however, differ markedly. Cytosolic and mitochondrial glutathione peroxidases (cGPXs or GPX1) reduce only soluble hydroperoxides, such as H2O2, and some organic hydroperoxides, such as hydroperoxy fatty acids, cumene hydroperoxide or t-butyl hydroperoxide. GPX1 and the phospholipid hydroperoxide glutathione peroxidase GPX4 (or PHGPX) are found in most tissues. GPX4 is located in both the cytosol and the membrane fraction. Furthermore, it can also directly reduce more complex lipids such as phosphatidylcholine hydroperoxides, fatty acid hydroperoxides and cholesterol hydroperoxides which are produced in peroxidized membranes and oxidized lipoproteins (32-35). GPX3 is directed to extracellular compartments and is excreted from various tissues in contact with body fluids. It reduces phospholipid hydroperoxides and contributes to extracellular antioxidant status in humans. However, the importance of its function is still questionable because of the low plasma concentration of glutathione (33).
GPX1 prevents cytotoxic peroxide-induced oxidative damage, lipid peroxidation and protein degradation. GPX4 is required for embryogenesis and male fertility. The exact function of GPX3 is still unknown and GPX2 might be an anti-inflammatory and anti-carcinogenic enzyme (36).
2. Regulation of synthesis
Exogenous selenium supply has been found to control the enzymatic activity of human GPX1. In a selenium-deficient environment, cells have about 5% of normal human GPX1 activity. However, GPX1 mRNA level is not affected by selenium level, which suggests that human GPX1gene is regulated post-transcriptionally by selenium (37,38).
Recently, it has been found that GPX1 is induced by etoposide, a topoisomerase II inhibitor, an apoptosis inducer and a p53 activator. DNA-binding assays have proved that p53 positively regulates an upstream promoter element of the GPX1 gene. This transactivation of GPX1 by p53 links the p53 signalling pathway to the antioxidant pathway (38). Moreover, analysis of the p53-induced apoptosis in a human colon cancer cell line showed that elevated p53 expression was associated with elevated GPX1 (38,39).
In addition, hyperhomocysteinemia is one of the risk factors for atherosclerotic vascular disease. Homocysteine has been reported to inhibit the expression of GPX1 and lead to an increase in reactive oxygen species that inactivated nitric oxide and promoted endothelial dysfunction. The overexpression of GPX1 rescued this homocysteine-induced endothelial dysfunction (40,41).
3. Cellular signalling
GPX1 counteracts hydroperoxide-modulated events, such as cytokine signalling and CD95-triggered apoptosis, to eliminate neoplastic cells (42,43), and GPX1 also has an important role in human immunodeficiency virus (HIV)-infection (44).
Glutathione peroxidases modulate NF-κB activation, which has been confirmed by the following considerations: inhibitors of cyclo-oxygenases and lipooxygenases inhibited activation of NF-κB, the activity of cyclo-oxygenases depends on the hydroperoxide tone which is regulated by glutathione peroxidases, NF-κB activation was inhibited in selenium-supplemented cells and facilitated in selenium deficiency. Moreover, experiments have implicated that GPX1 overexpression in human T47D carcinoma cells inhibited TNF-induced activation of NF-κB and modulated the phosphorylation pattern of hsp27 upon TNF treatment. However, GPX4 proved to be a more attractive candidate for silencing lipooxygenases and influencing cytokine signalling (45).
Apoptosis or programmed cell death plays an important role during embryonic development, in tissue remodelling, balancing carcinogenesis and host defense system. It can be induced in T cells by ROS, Fas antigen or TNF. Most of the studies of the GPX1 role in apoptosis were done in cells derived from lymphocytes. Enhanced GPX1 activity inhibited apoptosis induced by hydroperoxides. This fact was confirmed by supplementation of bovine renal epithelial cells with selenium or by overexpression of GPX1 construct in a myeloic cell line (33,46,47).
Finally, the role of GPX in HIV infection has been studied in detail. The replication of the HIV depends on the activation of NF-κB. Low levels of GSH and glutathione peroxidases in CD4+ cells enhance the level of hydroperoxides, leading to stimulation of apoptosis. The HIV-infected cells die in an apoptotic process. The apoptosis is efficiently inhibited by the anti-apoptotic Bcl-2 gene product, which has been shown to exert an antioxidant function. Similar results have been obtained with the cells overexpressing GPX1. Thus, GPX1 and Bcl-2 display analogous effects on an antioxidative event in the signalling cascade leading to inhibition of apoptosis, although by different mechanisms. GPX1 directly reduces hydroperoxides, whereas Bcl-2 prevents their formation. Based on these studies and studies involving GSH depletion, it was concluded that a reduction in GSH levels and GPX activity before infection could decrease spread of the virus due to apoptosis caused by oxidative microenvironment (48,49).
4. Pathophysiologic functions of enzymes
In case of head and neck cancers, GPX1 allelic loss has been shown to occur in histopathologically normal tissue adjacent to tumors, indicating that loss at this locus may be an early event in cancer evolution. GPX1 knockout mice have been created to study the consequent physiologic function of this enzyme. These mice grew and developed normally and did not show any histopathologies up to 15 months of age, indicating a limited role of GPX1 during normal development and under physiologic conditions (50).
However, when stressed with paraquat, GPX1(-,-) mice died faster than controls. Neurons from GPX1 knockout mice were also more sensitive to H2O2. Moreover, eye lenses from knockout mice were less able to recover than those of control mice when exposed to photochemical stress. GPX1 (-,-) mice expressed unchanged levels of GPX4, GPX3 and GPX2, which may particularly substitute for the GPX1 deficiency (50).
Human genetic alterations lead to asymptomatic state of decreased GPX activity, GSSG reduction or NADPH supply (33).
Chu et al. suggested a possible protective role of GPX2 against colon cancer. Such role has been shown from chromosomal mapping of the mouse GPX2 gene near the Ccs1 locus on chromosome 12, which contains a colon cancer susceptible gene. GPX2 mRNA levels were observed to be higher than in sensitive ICR/HA mice (31).
GPX3 has been first detected in plasma, yet its mRNA was predominantly found in epithelial cells of renal proximal tubules. Patients with renal diseases had very low GPX3 activity, including those undergoing renal dialysis. This decrease in GPX3 in plasma was not associated with selenium deficiency in patients. This fact represents the antioxidative protective role of this enzyme in the proximal tubules of the kidney (51,52).
In situ hybridization revealed that GPX4 is abundantly expressed in late spermatogenic cells. Oxidative GPX4 inactivation is obviously a crucial step in sperm maturation. The nuclear form of GPX4 is believed to contribute to chromatin condensation (53). GPX4 was shown to function as a structural protein in sperm heads, where it constitutes 50% of the protein (54). Some characteristic of different GPXs are assembled below in table 2.
Table 2. Characteristics of different GPXs
Conclusion
In conclusion, at least 7 types of glutathione peroxidases have been identified. Besides their intrinsic scavenger functions, precise localization of various GPXs in tissues argues in favor of specific roles of these enzymes. Each GPX could also function as a hydrogen peroxide sensor to regulate the H2O2 concentration. Interestingly, the specific cotranslational incorporation of Sec into GPXs has a dual effect in eukaryotes, such as recoding the UGA codon and stabilization of the mRNA in a selenium-dependent manner, which could be studied in the future for each type of GPX.
Due to the antioxidative and antimutagenic role of GPXs, there is a considerable interest in their therapeutic use as antioxidants. This may involve the use of naturally occurring antioxidants or completely synthetic molecules. Furthermore, overexpression of GPXs might protect various cells from oxidative stress. The activity of these enzymes might be elevated by adenoviruses (62), selenides, diselenides and ebselen as small-molecule GPX mimetics (63-65). The modelling of novel substances mimicking different GPXs and their transport to specific sites in anticancer therapy or its prevention are expected to become a trend in near future.