Introduction
Rheumatoid arthritis (RA) with a prevalence of 0.5% to 1% in the general population is the most common inflammatory systemic autoimmune disease. Characterized by chronic inflammation and consequent irreversible joint damage, but also damage to other organ systems, the disease considerably affects the patient’s quality of life, thus posing a major socioeconomic problem. Considering a recent concept according to which irreversible joint lesions occur in the first two years of the symptom onset, current therapeutic approach to RA implies aggressive anti-rheumatic therapy already in the first months of the disease. Due to the potentially toxic drugs that can cause severe side effects, a highly specific marker is necessary to identify RA patients before the onset of joint lesions, where the benefit of this therapy would overcome the potential risks. The American Rheumatism Association (ARA) criteria for RA diagnosis (1) that are primarily based on clinical indicators, except for rheumatoid factor (RF), are insufficiently sensitive, especially in this critical early stage of the disease. After discovery of the RF marker, which showed acceptable sensitivity but insufficient RA specificity, years have been spent on efforts to identify another, more specific antibodies. The majority of described antibodies have not shown diagnostic superiority regarding RF, until recent discovery of a family of antibodies directed to citrullinated epitopes on proteins/peptides(ACPA), which have been found to be a specific diagnostic as well as prognostic marker of RA.
Citrullination – a key process in the autoimmune response in RA
Today it is known that antibodies from ACPA family had long been identified in serum of RA patients, but insufficient knowledge of the structure of antigenic determinants resulted in tests of unsatisfactory diagnostic characteristics. The history of ACPA dates back to 1964 when Nienhuis and Mandema (4) using the method of indirect immunofluorescence (IIF), discovered antibodies in serum of RA patients that bound to the components of keratohyaline granules surrounding the nucleus of human oral cavity epithelial cells, giving characteristic perinuclear fluorescence. Regarding the fluorescence type, these antibodies were named antiperinuclear factor (APF). According to the results of studies that followed this discovery, APF antibodies are present in 49%–91% of RA patients, with 73%–99% specificity (3). However, despite high specificity, determination of these antibodies has not been accepted in daily routine for two reasons. The first is limited availability of appropriate substrate because epithelial cells differentiated enough to contain perinuclear factor can only be found in about 5% of donors. The second reason refers to unfavorable characteristics of the IIF method such as the lack of standardization and of experience required for correct interpretation.
In 1979, using the IIF method Young et al. (4) identified antibodies with target antigen located in filaments of the keratinized epithelial layer of rat esophagus. Assuming keratin as the target antigen, they named these antibodies antikeratin antibodies (AKA). AKA sensitivity for RA is 36%–59% and specificity 88%–99% (3). The autoimmune origin of these antibodies was proven by their binding to, at the time unknown, component in the keratinized layer of human epidermis. Several arguments have supported the hypothesis on the AKA and APF to belong to the same family: 1) target antigen for both antibodies is expressed in the epithelium of the same histologic type (rat esophageal epithelium, human epidermis and oral cavity epithelium belong to squamous epithelium with similar pathways of final differentiation); 2) their presence and titer correlate with each other, with RF and with the disease severity and activity; 3) both antibodies are mostly of IgG isotype; 4) both are highly specific for RA; 5) their prevalence is neither sex nor age dependent; and 6) both antibodies are present in the early stage of the disease and may precede the onset of symptoms. In 1993, Simon et al. (5) identified a 40kDa protein extracted from human epidermis and specifically marked by serum from RA patient on Western blot as the acidic/neutral isoform of filaggrin. Two years later, Sebbag et al. (6) found the IgG fraction purified from RA patient serum by immunoadsorption using the mentioned 40kDa protein to be reactive on both AKA and APF tests, demonstrating that these antibodies recognized the same antigen. Thus, the target antigen identified was filaggrin, a protein involved in the cytokeratin filament aggregation during epidermis keratinization. With this discovery, a new name for the antibodies has been introduced, including AKA and APF, i.e. anti-filaggrin antibodies (AFA). Filaggrin is synthesized in stratum granulosum as a phosphorylated precursor, profilaggrin (200–400 kDa), that accumulates in cytoplasmic organelles specific for keratinocytes, keratohyaline granules. During final differentiation of epidermal keratinocytes, profilaggrin molecules are dephosphorylated and proteolytically cleaved into functional filaggrin units. As filaggrin contains 10%–12% of the basic amino acid histidine, the question was raised of the origin of the acidic and neutral filaggrin isoform recognized by AFA. The basic filaggrin molecules released by the profilaggrin precursor proteolysis are post-translationally modified by the peptidylarginine deiminase (PAD) enzyme, whereby 20% of arginine residues are being deiminated (depending on Ca++ ions) to the citrulline amino acid (Figure 1).
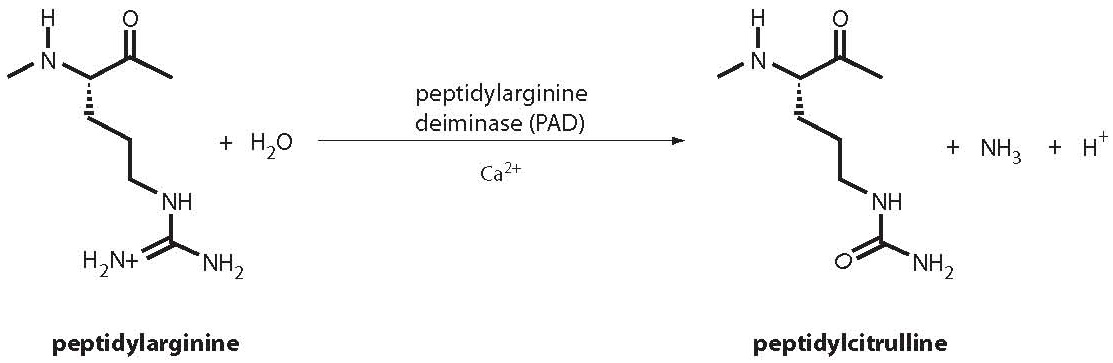
Figure 1. Peptidylarginine deimination to peptidylcitrulline by Ca++ ion-dependent peptidylarginine deiminase (PAD).
By the procedure of citrullination, the basic isoform transforms to the neutral or acidic filaggrin isoform, which has lower affinity for cytokeratin filaments. In 1998, Schellekens et al. (7) successfully identified citrulline as the main antigen determinant using citrullinated peptides synthesized towards the COOH-terminal end of the profilaggrin molecule. Only a year later, this discovery was confirmed by Girbal-Neuhauser et al. (8), showing the RA-specific antibodies to recognize recombinant filaggrin fragments after enzymatic deimination in vitro.
Citrullinated proteins in rheumatoid synovial tissue
Due to the fact that (pro)filaggrin expression is limited exclusively to squamous epithelium and is not present in synovial tissue, it is assumed that deimination of different proteins in inflamed synovial tissue activates immune reaction, and citrullinated (pro)filaggrin is merely a cross-reactive substrate (9). In favor of this hypothesis speaks the result of Masson-Bessiere et al. (10) who discovered a 7.5-fold higher AFA ratio in total IgG concentration in the extract of rheumatoid synovial tissue than in the respective serum. This result points to a conclusion that AFA-secreting plasma cells are present in synovial tissue of RA patients and the locally secreted antibodies are then being diffused into the circulation. The same authors suggest deiminated isoforms of a- and β-fibrin chains, deposited in rheumatoid synovial membrane, as the main target antigens for AFA (11). However, a recent study has shown that the presence of citrullinated fibrin is not specific for rheumatoid synovial tissue, but is also found in inflamed synovial tissue in spondyloarthropathies (psoriatic arthritis, ankylosing spondylitis and reactive arthritis) and osteoarthritis. Therefore, fibrin deimination is a nonspecific phenomenon associated with synovitis which need not necessarily induce autoimmune response with the generation of AFA antibodies (12). Human type I collagen (13), vimentin (14) and fibronectin (15) have been identified among other citrullinated synovial proteins. Furthermore, Kinloch et al. (16) demonstrated reactivity of 46% of RA sera with in vitrocitrullinated a-enolase from promyelocytic cell line lysate that is differentiated into cells of monocyte and granulocyte phenotype, which prevail in inflamed synovial tissue. Although the presence of a-enolase has been demonstrated in synovial tissue, its in vivocitrullination has not yet been confirmed. Bearing in mind that citrulline bound to neutral amino acids like glycine, serine or threonine is critical for AFA binding, Pratesi et al. (17) investigated the hypothesis whether deimination of Epstein-Barr nuclear antigen 1 (EBNA-1) that contains 6 repeating arginine-glycine sequences and is often related to RA, can generate epitopes recognizable by AFA. Antibodies specific for this EBNA-1 sequence have been detected in 50% of RA sera and in <5% of sera from normal controls, but the presence of deiminated EBNA-1 in rheumatoid synovial tissue has not been proven. Finally, although the presence of citrullinated histones in inflamed synovial tissue has not yet been proven, these proteins do not seem as negligible potential antigens, since their citrullination has been described ex vivo during granulocyte apoptosis (18). A great number of granulocytes with a life span of some 3 days are present in inflamed synovial tissue, and many of them will undergo apoptosis at one time. It is accompanied by citrullination of histones, which may trigger local immune response.
If protein deimination occurs locally, in synovial tissue, the PAD enzyme must be present. This enzyme is ubiquitous in mammalian tissues and 5 isoforms with tissue-specific expression have been identified to date. PAD2 and PAD4 are, among other tissues, also expressed in hematopoietic cells (22), and are therefore of special interest because they infiltrate inflamed rheumatoid synovial tissue. Chapuy-Regaud et al. (23) demonstrated expression of these two isoforms in synovial membrane of RA patients.
The mechanism of RA-specific immune response to citrullinated antigens
PAD enzyme isoforms are present in different tissues, so the process of citrullination is part of the normal physiologic processessuch as already described final differentiation of epidermal keratinocytes or of myelin sheath during central nervous system development (21). Besides rheumatoid synovial tissue, citrullinated protein expression was also observed in synovial tissue of patients suffering from osteoarthritis, reactive arthritis and other arthropathies (22). Furthermore, citrullinated proteins were detected in neurodegenerative diseases like Alzheimer’s disease (23), multiple sclerosis (24), but also in muscles of patients suffering from polymyositis, in the intestine of patients with ulcerative colitis and Crohn’s disease and chronic tonsillitis (25). It appears that the link between the process of citrullination and these diseases lies in inflammation. However, in these cases the presence of citrullinated antigen is not connected with simultaneous presence of autoantibodies, which means that citrullinated proteins, even in inflammatory environment, are not sufficient to activate specific immune reaction. How then to explain the presence of these highly specific autoantibodies in RA? There are three suggested mechanisms that might be involved in RA-specific immune response to ubiquitous antigens like citrullinated proteins: 1) it is known that immune response is defined not only by the mere presence but also by the quantity of the antigen present, which would mean that there is an over-expression of citrullinated antigens in rheumatoid synovium; 2) abnormal humoral immune response to commonly present citrullinated proteins in inflamed tissue; and 3) presence of RA-specific citrullinated epitopes (26). It seems that certain genetic factors could be responsible for the first two mechanisms. Suzuki et al. (27) found an association between polymorphisms of the PADI4 gene encoding PAD4 enzyme and RA in the Japanese population. These polymorphisms were found to result in a mRNA of threefold stability, pointing to enhanced PAD4 enzyme synthesis in persons with this haplotype. This enzyme is present in the nuclei of granulocytes and monocytes that grossly infiltrate inflamed rheumatoid synovial tissue; so, the elevated level of PAD4 enzyme in these cells will have a cumulative effect on the final quantity of citrullinated proteins. Thus, a high quantity of citrullinated antigenes is presented to the immune system, which can lead to the loss of tolerance to own antigens. However, the association of PADI4 polymorphisms and RA should be considered with caution, since it has not been confirmed in other populations (28). Another mechanism that could be responsible for excess citrullinated antigen expression is inefficient clearing of dying cells. The intercellular Ca++ ion concentration is too low for PAD enzyme activation, so it remains inactive. It is only activated when Ca++ ion homeostasis has been disturbed, e.g., upon final epithelial cell differentiation. During cell death, the integrity of cellular membrane is impaired, which can have a dual effect, i.e. Ca++ ion influx from extracellular space and activation of intracellular PAD enzyme or PAD enzyme efflux into extracellular space with consequential massive protein citrullination. In conditions predominated by inefficient dead cell removal (due to massive cell death or inefficient mechanism of their elimination) such as RA, a large quantity of citrullinated proteins will be exposed to the immune system, which will recognize them as foreign antigens (26). The next genetic factor that can affect abnormal humoral response to commonly present citrullinated proteins are HLA-DR alleles associated with an increased risk of RA, primarily HLA-DRB1*0401 and HLA-DRB1*0404. Namely, a recent study has shown that vimentin derived citrullinated proteins but not those having arginine on the same locus, bind to HLA- DRB1*0401 molecule with high affinity (29). This interaction may underlay the citrulline-specific immune response, because in an experiment with HLA-DRB1*0401 transgenic mice the same authors proved that stimulation with citrullinated peptides, but not with the arginine variant of peptide, stimulated T lymphocyte proliferation and activation. However, the absence of ACPA antibodies in HLA-DRB1*0401 positive patients with spondyloarthritis and psoriatic arthritis, with the presence of deiminated fibrin detected in their synovial membrane, suggests that the haplotype associated with an increased risk of RA alone is not sufficient for ACPA-specific immune response (12). Furthermore, polymorphism of the gene for cytokines that play an important role in inflammatory process characteristic of RA is a potential genetic factor that may be involved in RA-specific immune response to citrullinated proteins. One of these polymorphisms is -2849 A/G in the promotor region of the IL-10 gene responsible for enhanced production of IL-10. Besides numerous anti-inflammatory effects, this interleukin also exerts proinflammatory action by stimulating B lymphocyte proliferation and differentiation with consequential antibody production. It has been shown that ACPA positive RA patients with a haplotype that results in high IL-10 concentration have higher antibody titer and more severe erosive lesions in comparison with ACPA positive RA patients without this haplotype (30). A higher locally formed antibody titer also leads to higher production of immunocomplexes, which bind via Fcg receptors to macrophages that are then activated and secret an increased amount of proinflammatory cytokines. Different polymorphisms of cytokines and their receptors have been associated with RA (31). These genetic factors cause the release of a higher amount of cytokines after stimulation or cause higher cell sensitivity to cytokines. As cytokines are the key molecules in inflammatory response that includes influx of a great number of inflammatory cells from the circulation and their activation, this continues the cycle that leads to chronic inflammation characteristic of RA. Besides the mentioned genetic factors that can contribute to RA-specific immune response to ubiquitous citrullinated proteins, the hypothesis on the existence of RA-specific intracellular citrullinated epitopes has currently been increasingly adopted. Intracellular citrullinated proteins which, unlike extracellular ones, are RA-specific, have been detected in rheumatoid synovium by immunohistochemical method using a mouse monoclonal antibody. These proteins show colocalization with PAD2 enzyme, which is significantly more expressed in RA synovial tissue than in synovial tissue of normal controls. Furthermore, correlation of intracellular citrullinated protein expression with local and systemic level of ACPA antibodies has been established (32). Biochemical identity of these deiminated intracellular proteins is still in the phase of research.
Methods of ACPA determination and their clinical significance
Synthetic citrullinated cyclic peptides or citrullinated proteins the presence of which has been demonstrated in synovial tissue extract are used in enzyme-linked immunosorbent assays (ELISA) for determination of these antibodies. On choosing protein substrate, those rich in arginine, such as fibrinogen and vimentin, are preferred. After in vitro citrullination, these proteins will have more different epitopes that constitute citrulline, but also neighboring amino acids, thus theoretically increasing the sensitivity to polyclonal immune response as compared with synthetic cyclic peptides. On the other hand, however, the use of protein as substrate has some disadvantages such as the problem of standardization of in vitro enzyme citrullination and the possible serum reactivity with other non-citrullinated epitopes.
Antibodies to cyclic citrullinated peptides (anti-CCP)
Identification of filaggrin as a target molecule recognized by RA-specific AKA and APF antibodies has enabled a new methodological approach to detection of these RA-specific antibodies. Immunoblot methods were the first methods employed, then also ELISAs were used with filaggrin extracted from human epidermis as antigen substrate (33,34). However, it turned out almost impossible to prepare a standardized substrate with filaggrin used as antigen. The reason is the fact that almost 30%–40% of amino acid residues on the filaggrin molecule are changeable and citrullination of arginine residues is only partial, so these isoforms are characterized by great charge heterogeneity. This problem also applies to in vitro enzymatic deimination of bacterially expressed recombinant filaggrin fragments. Contrary to heterogeneous natural substrate, synthetic peptides carry more characteristics of an ideal antigenic substrate. Peptides of predefined sequence can be synthesized in large amounts with high purity level and their small size reduces the possibility of nonspecific interactions that occur with other serum components. Still, the use of linear synthetic peptides (synthesized according to profilaggrin molecule) as a substrate in ELISAs proved disappointing because satisfactory sensitivity could only be achieved with the use of several different synthetic peptides, which was technically too demanding (35). One of the problems with the use of linear peptides as substrate is that adsorption of all peptides on the wells of ELISA microtiter plates cannot be achieved by the standard procedure of passive adsorption. Partial solution of this problem is covalent peptide binding to a solid carrier. Another problem encountered on the application of relatively short (linear) peptides is that they generally do not assume stable conformation in a solution and therefore only a fraction of the peptides assumes conformation comparable to that of the original binding site on the protein antigen. This effect named “conformational dilution” has a direct unfavorable effect on the antibody affinity for peptide despite the fact that peptide sequence spans the complete antigen determinant. The way to solve this problem is to enable the peptides bound to microplate wells to assume conformation that favors antibody binding, i.e. which will imitate conformation of the original binding site on the protein as close as possible. Having in mind that peptides often assume β-conformation in Ag-At complex and the fact that the cystine-bridged cyclic peptides imitate β-conformation of the original antigen determinant and enhance the antibody binding affinity, Schellekens et al. (35) used cyclic peptides formed by substitution of serine residues with cysteine which is then oxidized into cystine. Thus the first generation of ELISAs with cyclic citrullinated peptides (CCP1) as antigenic substrate were created. Median sensitivity calculated from studies using CCP1 test was 54% (ranging from 41% to up to 68%) and median specificity 97% (ranging from 90% to up to 99%) (36). Several years later, test sensitivity was improved in a way that a library of citrullinated peptides was searched using pool serum of RA patients, which resulted in identification of numerous high-reactive peptides that are currently used in second generation ELISAs (CCP2) (37). In 2005, third generation of the anti-CCP test (CCP3) appeared on the market, including additional citrullinated epitopes and a test version with simultaneous detection of IgG and IgA anti-CCP antibodies (CCP 3.1), which should improve test sensitivity. Although some authors (38,39) found improvement of sensitivity by about 5% in relation to CCP2, results of other studies showed an equal diagnostic efficiency of CCP2 and CCP3 tests (40,41).
Diagnostic accuracy of anti-CCP2 test in RA
Since 2003, a number of studies assessing diagnostic accuracy of anti-CCP2 test in RA have been published. The results show significant variability, which is attributed to differences in the characteristics of RA patient groups on the one hand and heterogeneity of control groups on the other hand. The sensitivity of anti-CCP2 in patients with advanced RA stage spans from 64% to even 96%, while in patients with early RA stage or those with undifferentiated arthritis it ranges from 14.4% to 83.5% (42). The choice of control groups influences test specificity. Healthy subjects have quite frequently been included in control groups, thus overestimating test specificity. Inclusion of patients with the same disease in control group has the same effect. The real diagnostic characteristics of a test can only be obtained in a study including patients that represent the real population in which the test will be used in clinical routine. Therefore, taking into consideration only studies where control groups corresponded best to this presumption, the specificity of anti-CCP2 for RA ranges from 90.4% to 97.3% (42). In addition to RA, anti-CCP antibodies have also been found in 9% of patients with systemic lupus erythematosus, 5% of patients with Sjögren’s syndrome, 8% of those with psoriatic arthritis and 2%–5% of patients suffering from juvenile idiopathic arthritis (36).
Predictive value of anti-CCP2 test for RA development
Rantapaa-Dahlqvist et al. established the presence of anti-CCP antibodies, as in case of RF, many years before the initial RA symptoms. Retrospective analysis of donor blood had identified 83 patients that subsequently developed RA. Anti-CCP antibodies are a predictive marker that is positive for 1.5 years before the first visit to physician. During the last 9 years, the sensitivity of anti-CCP antibodies has shown an increasing tendency from 4% to 52% (43). In a subgroup of the same cohort, anti-CCP antibodies showed a higher predictive value than RF and HLA-DRB1 antigens of the locus for RA development, with odds ratio (OR) of 15.9 two years before the onset of symptoms (44). Predictive value of anti-CCP antibodies for RA was also confirmed in longitudinal studies in cohorts of subjects presenting with nonspecific early inflammatory arthritis (undifferentiated arthritis). In the study by van Gaalen et al., at one year ARA criteria for RA diagnosis were satisfied by 75% of patients that initially were anti-CCP positive, and at 3 years the percentage increased to even 93% (OR 38.6 vs. 9.8 for RF). Among patients that initially were anti-CCP negative, only 25% were diagnosed with RA at 3 years (45). In another study, patients suffering from synovitis for a period of ≤3 months were monitored for 18 months. A combination of anti-CCP and RF was found to have a positive predictive value (PPV) of 58% and negative predictive value (NPV) of 88% for RA diagnosis (46). These results suggest that the occurrence of anti-CCP, like RF, is part of a very early process in the development of RA. Clinical significance of the predictive value of anti-CCP antibodies for RA development lies primarily in reducing the time elapsed from the first synovitis symptom onset to therapeutic intervention.
Prognostic value of anti-CCP2 test in RA
The use of first generation anti-CCP assays pointed to the potential of these antibodies in predicting erosive progression of RA. There are an increasing number of studies confirming it with the use of second generation assays. In the scope of a Swedish study (TIRA project), 242 patients with early stage RA (disease duration ≤1 year) were followed-up for 3 years by regular measurement of laboratory (erythrocyte sedimentation rate (ESR) and C-reactive protein (CRP)) and clinical indicators of disease activity (28 joint disease activity score (DAS28); swollen joint count; physician’s general assessment of disease activity; assessment of functional ability by the Health Assessment Questionnaire (HAQ); RF values (of IgM and IgA isotypes) and anti-CCP values. Although the diagnostic sensitivity of anti-CCP and RF was almost equal, positive anti-CCP antibodies at diagnosis proved to be better predictor of disease activity in the next 3 years. Anti-CCP positive status remained almost unchanged during 3 years irrespective of the antirheumatic therapy applied (47). In another study, Forslind et al. examined predictive value of several factors including anti-CCP antibodies for radiologically measured progression of articular damage using the method of Larsen in 379 patients with early RA during 2 years. Patients with positive anti-CCP antibodies had a significantly greater articular damage both at the beginning and at the end of monitoring than those with negative anti-CCP antibodies. The same tendency in relation to anti-CCP status was observed in laboratory and clinical indicators of disease activity (48). Logistic regression analysis pointed to the grade of articular damage according to Larsen, anti-CCP and the ESR measured initially as the most important variables in predicting the progression of articular damage. This model enables correct classification in terms of damage progression in 75% of cases. By monitoring 279 patients with early stage of RA, Rönnelind et al. confirmed the presence of anti-CCP antibodies initially to predict unfavorable course of the disease in the next 5 years and more progressive joint damage in 2 years (49). It should be noted that the results of this study have revealed that changes in anti-CCP antibody concentration do not reflect disease activity. In a recently published study, anti-CCP, IgM-RF, ESR and female sex were found to be independent predictors of articular damage progression over 10 years, with positive anti-CCP antibodies as the strongest predictor (50).
The predictive value of anti-CCP antibodies complemented with other variables such as ESR, radiologically assessed joint damage, RA-risk HLA-alleles or RF allows for very early identification of patients at a high risk of developing progressive erosive arthritis, where aggressive antirheumatic therapy is indicated.
Antibodies to citrullinated vimentin (anti-CV) and modified citrullinated vimentin (anti-MCV)
More than a decade ago, Despres et al. (51) described new antibodies in serum of RA patient, which they named Sa antibodies after family name of the patient where it was first identified (Savoie). Sa antibodies recognize protein band of 50 kDa on the human spleen extract immunoblot and 2 bands (50 and 55 kDa) on the human placenta extract immunoblot. These antibodies are present in 21%–43% of RA patients with a very high specificity of 92%–100% (52). In 2004, Vossenaar et al. (14) discovered several peptide sequences matching vimentin sequences by sequencing Sa gene purified from placenta extract. Using Western blot analysis, the same authors confirmed anti-Sa antibody binding to citrullinated recombinant vimentin but not to non-modified vimentin, suggesting the presence of citrulline to be essential for Sa autoantigenicity. With this discovery, Sa antibodies identified as antibodies to citrullinated vimentin (anti-CV) were included in the large family of antibodies to citrullinated proteins. Polymerized vimentin molecules constitute a structure of intermediary filaments, the main component of cytoskeleton in mesenchymal cells (chondrocytes, synovial fibroblasts) and macrophages, thus its presence in synovial tissue is expected. Intermediary filaments together with actin microfilaments and microtubules constitute a network responsible for mechanical cell integrity and are involved in critical processes like mitosis and motility. Polymerization of free vimentin subunits into filaments is a reversible process where dephosphorylation occurs (53). Vimentin is usually not found in citrullinated form in vivo, but deimination of arginine residues on vimentin molecule has been demonstrated to occur in macrophages during apoptosis (54). As in vitro deimination of the amino-terminal domain of the vimentin molecule is known to cause instability of the intermediary filament network, it has been postulated that vimentin citrullination in vivo might be involved in cytoskeletal collapse during apoptosis, when vimentin filaments are being deposited into perinuclear aggregates. In conditions accompanied by inadequate apoptotic material clearance, as in RA, citrullinated vimentin is exposed to immunocompetent cells that react by the production of autoantibodies. However, as citrullinationper se is insufficient to trigger such an immune reaction, it has been postulated that inflammatory microenvironment predominated by reactive oxygen and nitrogen radicals can induce gene mutations for vimentin and thus create new antigens in synovial fibroblasts and macrophages. Vimentin isoforms were purified from fibroblasts and synovial fluid from RA patients, characterized by LC/MS spectroscopy, and mutations and citrullination were identified. Recombinant mutated vimentin isoforms were expressed in E. coli, whereas purified protein was citrullinated in vitro and used as antigen in ELISA for determination of IgG antibodies to mutated citrullinated vimentin (anti-MCV).
Diagnostic accuracy and prognostic value of anti-MCV antibodies in RA
Despite expectations that the use of citrullinated protein as target antigen instead of synthetic peptides could increase test sensitivity for ACPA antibody detection, results of the studies reported to date point to comparable diagnostic accuracy of anti-MCV and anti-CCP2 tests. Dejaco et al. compared 164 RA patients and control group of patients suffering from other inflammatory and non-inflammatory rheumatic diseases, and found 69.5% sensitivity and 90.8% specificity for anti-MCV test vs. 70.1% sensitivity and 98.7% specificity for anti-CCP2 test (55). In another study, anti-MCV antibodies showed sensitivity higher by 9% and 4% in comparison with anti-CCP2 antibodies and RF, respectively. However, diagnostic accuracy according to ROC analysis was somewhat but not significantly lower than for anti-CCP2. Although these antibodies were in significant correlation, pointing to many common epitopes, the presence of only one antibody in about 12% of patients suggests that citrullinated antigens in either test contain unique epitopes that are only recognized by one or the other antibody. Therefore, the authors suggest combined determination of anti-CCP2 and anti-MCV in order to improve diagnostic efficiency (56). Mathsson et al. determined anti-CCP and anti-MCV antibodies in 273 patients with early RA at the time of symptom onset, at three months and at one year; in 72 patients anti-CCP and anti-MCV antibody determination was continued at 2, 3 and 5 years (57). In this study, the sensitivity of anti-MCV antibodies significantly exceeded (70.7%) that of anti-CCP2 (57.9%), with comparable specificity (95% vs. 96%). However, it should be noted that diagnostic accuracy refers to comparison of patients with early RA and healthy subjects. Positive anti-MCV antibodies at first symptom onset turned out to be as good predictor of progressive erosive arthritis as anti-CCP antibodies. It seems that anti-MCV antibodies are a more sensitive indicator of disease activity. Namely, changes in the concentration of anti-MCV antibodies during one year yielded higher correlation with changes in laboratory (ESR and CRP) and clinical indicators of disease activity (DAS28, HAQ, doctor’s assessment of disease activity, swollen joint count). In a recent study, Innala et al. confirmed higher association of anti-MCV antibodies with more severe course of the disease in comparison with anti-CCP antibodies determined by second and third generation tests, with almost equal predictive value for progression of radiologically assessed articular damage (58).
Based on the results of the studies reported to date, it seems that anti-MCV antibodies failed to make any significant diagnostic contribution, however, their determination in early RA is clinically relevant for their prognostic value for a more severe course of the disease.
Conclusion
Modern therapeutic approach to RA that assumes early introduction of aggressive antirheumatics to prevent irreversible articular damage poses the need of early verification of the diagnosis and prognosis of the course of disease. Current diagnostic approach based on clinical parameters and RF as the only serologic marker has proved inadequately sensitive and specific in the critical, early stage of the disease. The discovery of citrulline as the main determinant of target antigens of highly specific antibodies in RA has marked the beginning of a new era in the diagnosis of RA. The use of synthetic cyclic citrullinated peptides (CCP) as antigens in ELISAs has increased the sensitivity of these antibodies for RA. Furthermore, these highly specific antibodies have turned out to be a valuable prognostic marker. The ubiquity of citrullinated proteins in physiological and pathological conditions as well as the presence of antibodies to citrullinated proteins many years before the first RA symptom onset suggest that citrullination per se is not sufficient for RA development. Therefore, understanding of the mechanisms of RA-specific immune response to citrullinated proteins has been in the focus of research in the last years. One of the most important goals in these researches is identification of citrullinated proteins as potential antigens in inflamed synovial tissue. Up to now, numerous proteins have been identified as potential native antigens, such as citrullinated vimentin, fibrin and histones. According to latest findings, it seems that distinct citrullinated epitopes rather than mere presence of citrullinated proteins are responsible for the induction of immune response. Therefore, defining of these epitopes remains as the major challenge in forthcoming researches.