Introduction
5’-Nucleotidase (5’-NT) is an intrinsic membrane glycoprotein, present as an ectoenzyme in a wide variety of mammalian cells. 5’-NT hydrolyzes 5’-nucleotides to their corresponding nucleosides (1). It is measured as an indicator of liver damage resulting primarily from interference with the secretion of bile (2). Enzyme activity is elevated in liver diseases including liver cirrhosis, chronic alcoholism, neoplasms of the liver and bile ducts, benign biliary disease, but reaches its highest level in the presence of biliary stasis (3,4). The diagnostic value of 5’-NT has been shown to be superior to other liver enzymes, especially in liver metastasis. Raised levels of 5’-NT activities are found in 92% of patients with obstructive jaundice, 70% of patients with parenchymal liver disease and 81% of patients with hepatic metastasis (5,6). It is also reported that serum 5’-NT is clinically useful for differential diagnosis of hepatobiliary and osseous diseases, the enzyme activity being only increased in hepatobiliary disease (7).
Although determination of either oxidants or antioxidant components alone may give information about oxidative stress, determination of oxidants along with antioxidants is more useful in this context (8,9). Studies suggest that evidence of oxygen free radical is found early in the development of fibrosis and cirrhosis of the liver (10). Patients suffering from liver disease either due to non-alcohol or excessive alcohol intake show depletion of antioxidants such as glutathione (GSH) and superoxide dismutase (SOD), and increased concentration of the products of lipid peroxidation such as malondialdehyde (MDA) (11-19). Hepatic fibrogenesis in alcohol liver cirrhosis is an intricate process, which appears to involve metabolic products of ethanol oxidation, cytochrome P450 induction, enhanced oxidative stress, depletion of antioxidant defenses, lipid peroxidation, generation of aldehydic products, effects of mitogenic and fibrogenic cytokines, and complex interactions between liver parenchymal and non-parenchymal cells with hepatic stellate cells (Ito cells, hepatic lipocyte or fat storing cell) are now recognized as the primary source of extracellular matrix (20,21).
There is evidence that the diseased liver of patients with cholestatic liver disease is exposed to oxidative stress associated with increased lipid peroxidation involving intraorgan generation of reactive oxygen species (ROS), possibly mediated by endotoxins, bile acids and accumulation of degradation products of lipid peroxidation, such as lipid peroxides and MDA (20,22). Alcohol promotes the generation of ROS and/or interferes with the body’s normal defense mechanism against these compounds (23). Studies suggest that evidence of oxygen free radicals is also found early in the development of fibrosis and cirrhosis of the liver (10).
The present study was therefore undertaken tomeasure the 5’-NT enzyme activity in liver cirrhosis and alcohol consumers. Oxidative stress, antioxidants and their association with 5’-NT were also investigated.
Materials and methods
Subjects
This study was conducted in three groups of 25 subjects each, age range 35-70 years. There were 13 males and 12 females in each group.
Group I (control group): healthy individuals, age range 35-70 years (median 55, IQR 45-60) having no history of liver disease or alcohol intake in the past or present.
Group II (alcohol consumer group): age range 35-70 years (median 55, IQR 45-60) with a history of 10 years or more of alcohol intake without any history of liver disease or other hepatobiliary disorders. Alcohol consumers were taking 6250-7500 mg of alcohol/day during the study. Alcohol consumers were recruited by personal contact from known alcohol consumers.
Group III (cirrhotic patient group): age range 35-70 years (median 55, IQR 45-60) diagnosed with liver cirrhosis and with a history of jaundice, alcohol intake or hepatotoxic drug intake. Patients were recruited from the following hospitals in Mangalore: District Wenlock Hospital; K.M.C Hospital, Attavar; K.M.C. Hospital, Ambedkar Circle; A.J. Institute of Medical Sciences, Kuntikana; and Yenepoya Hospital, Kodialbail.
On selecting subjects for test and control groups, care was taken to eliminate subjects with habits like smoking, tobacco chewing, and those with chronic inflammatory diseases like tuberculosis, rheumatoid arthritis, diabetes mellitus and malignancy, all of which play a vital role in contributing to oxidative stress injury. Exclusion criteria were any bone diseases or any other clinical conditions that might be involved in raised 5’-NT activity.
Approval to carry out these studies in humans was obtained from the Institutional Clinical Ethics Committee of Kasturba Medical College, Mangalore, India and their guidelines were followed during the study.
Sample collection
Two mL of venous blood was collected in a plain vial for serum separation for the analysis of 5’-NT. Five mL of venous blood was collected in separate EDTA containers from the median cubital vein or basilic vein under strictly aseptic precautions. Of these, 0.2 mL of whole blood was used for glutathione estimation; the rest of the sample was centrifuged at 3000 rpm for 10 minutes within 3 h of collection. Plasma was discarded. The sediment was used to prepare erythrocyte suspension and used for the assay of MDA and SOD.
Methods
Preparation of erythrocyte suspension
The sediment separated was washed thrice with 0.9% cold normal saline and then suspended in an equal volume of the same saline solution. It was stored as 50% cell suspension at 4-5 °C for 24 h and thenused for the assay of MDA and SOD.
5’-nucleotidase determination
Serum 5’-NT enzyme was estimated using the method reported elsewhere (5). Two test tubes were set up as follows:
- Total activity: 0.2 mL of serum were added, 0.1 mL of 0.02M manganous sulfate and 1.5 mL of 40 mM, pH 7.5 barbitone buffer.
- Non-specific alkaline phosphatase activity:0.2 mL of serum were added, 0.1 mL of 0.02 M manganous sulfate, 1.3 mL of 40 mM, pH 7.5 barbitone buffer and 0.2 mL of 0.1M nickel chloride.
- Both test tubes warmed to 37 ºC, then 0.2 mL 10mM adenosine 5’-phosphate were added to each test tube and incubated at 37 ºC for 30 minutes. Then 2 mL of 10% trichloroacetic acid (TCA) were added, mixed well and left to stand briefly, then centrifuged. Two mLof supernatant were taken (=0.1 mL serum) for estimation of inorganic phosphorus. For the blank and standard 1 mL of water and 1 mL of the phosphate standard (stock solution containing 6 mmol/L) were taken for use, each with 1 mL of TCA added. To all four tubes 3 mL 2M, pH 4.0 acetate buffer, 0.5 mL of 5% ammonium molybdate and 0.5 mL metol (2 g metol and 10g sodium sulfite in water made up to 100 mL) mixed, allowed to stand for 10 minutes and then read using a red filter or at 680 nm. Percent coefficient of variation (% CV) of this method was 1.96.
Malondialdehyde (MDA)
Red cell lipid peroxidation was studied as thiobarbituric acid (TBA) reaction products. The method of Stocks and Dormandy was followed with certain modifications (24). One mL of erythrocyte suspension was added to 8.5 mL of 0.9% normal saline and mixed well. Then 0.5 mL of 0.44 M H2O2 was added. From the above mixture, 2.5 mL of aliquot was immediately taken, to which 1 mL of 28% TCA in 0.1 M sodium metaarsenite was added. This was mixed well and allowed to stand for 10 minutes, after which it was centrifuged. Three mL of the supernatant were then taken to which 1 mL of 1% TBA in 50 mM NaoH was added. This was then kept in a boiling water bath for 15 minutes and immediately cooled under tap water. The pink chromogen was read at 535 nm in a spectrophotometer. Values were expressed as nanomoles of MDA formed per dL of RBC, taking the molar extinction coefficient as 1.56 x 105. The % CV of this method was 1.88.
Glutathione (GSH)
Whole blood glutathione level was measured by the method of Beutler et al. (25). To 0.2 mL of whole blood, 1.8 mL of distilled water was added and mixed; 3 mL of precipitating solution was added, mixed and allowed to stand for 10 min at room temperature. This mixture was then centrifuged. To 2 mL of the supernatant, 8 mL of phosphate solution (0.4M) and 1 mL of DTNB reagent were added. The absorbance was read at 412 nm. The % CV of this method was 1.91.
Superoxide dismutase (SOD)
The method of Fridovich was followed on estimation of SOD (26). This method is based on the inhibition of nitroblue tetrazolium (NBT) reduction by superoxide radicals, generated by the illumination of riboflavin in the presence of oxygen and electron donor. Methionine was used as a basis for the assay of SOD. The % CV of this method was 1.75.
Preparation of hemolysate
It was done by the method of McCord and Fridovich (27). To 1 mL of erythrocytes (washed with 0.9% normal saline), 1 mL of deionized water was added to lyse the cells. To this 0.5 mL of distilled ethanol were added, followed by 0.3 mL of chloroform, mixed well and allowed to stand for 15 min. Now 0.2 mL of H2O were added and centrifuged at 4 ºC. The supernatant contained SOD activity and was used for the assay of SOD; after dilution with potassium phosphate buffer (pH 7.8, 0.05 M) 0.1 mL of hemolysate was diluted with 1.9 mL of potassium phosphate buffer. It was the final diluted hemolysate that was used in the procedure described below.
Four test tubes were taken and labeled as Test, Control, Test blank and Control blank. To the Test tube, 2.9 mL of reaction mixture with NBT containing 149 mg of methionine, 4.93 mL of NBT (1 mg/mL), 0.63 mL of riboflavin (1 mg/mL) were added and made up to 100 mL with potassium phosphate buffer (pH 7.8/0.05M), and 0.1 mL of diluted hemolysate was added. To the ‘Test blank’ tube, 2.9 mL of the same reaction mixture without NBT and 0.1 mL of diluted hemolysate were added.
To the Control test tube, 2.9 mL of the same reaction mixture with NBT and 0.1 mL of potassium phosphatebuffer (pH 7.8/0.05M) were added.
To the Control blank tube, 2.9 mL of the same reaction mixture without NBT and 0.1 mL of potassium phosphate buffer (pH 7.8/0.05M) were added. Each of these mixtures was now put into a 10-mL beaker. The beakers were kept in an aluminum foil lined box fitted with a 15 W fluorescent lamp for 10 minutes. The absorbance was read at 560 nm in a spectrophotometer for all the four beakers.
Determination of hemoglobin
The hemoglobin content of erythrocytes was determined by the cyanmethemoglobin method (28). Hemoglobin was treated with a reagent containing potassium ferricyanide, potassium cyanide and potassium dihydrogen phosphate (Drabkins Reagent). The ferricyanide oxidizes hemoglobin to methemoglobin, which is converted to cyanmethemoglobin by cyanide. The absorbance was measured at 540 nm in a spectrophotometer. The % CV of this method was 1.65.
Statistical analysis
All biochemical parameters were compared between different groups using Kruskal Wallis test. The statistical software SPSS Version 11 (statistical package for social sciences) was used for this purpose. The value of P<0.05 was considered as significant.
Results
In this study, serum 5’-NT activity was highest in the cirrhotic group, relatively high but less than cirrhotic and more than control in alcohol consumer group, and lowest in control group (Table 1). There was statistically significant difference in 5’-NT activity between three groups (P < 0.05). The highest 5’-NT activity recorded in the group of cirrhotic patients appeared to be indicative of greater sensitivity for intrahepatic obstruction or liver cell damage (2). Because it is a plentiful primary liver enzyme, 5’-NT may be more readily influenced by minute areas of obstruction.
The RBC MDA levels were highest in the group of cirrhotic patients, as shown in Table 1. MDA level was relatively lower in the group of alcohol consumers and was lowest in controls. There was statistically significant difference in MDA level between three groups (P < 0.05).
Blood glutathione levels were highest in the control group, relatively lower in alcohol consumer group and lowest in cirrhotic patient group, as shown in Table 1. There was statistically significant difference in glutathione concentration between three groups (P < 0.05).
Table 1. Concentrations of 5’-nucleotidase (5’-NT), malondialdehyde (MDA), glutathione (GSH) and superoxide dismutase (SOD) in control, alcohol consumer and cirrhotic patient groups.
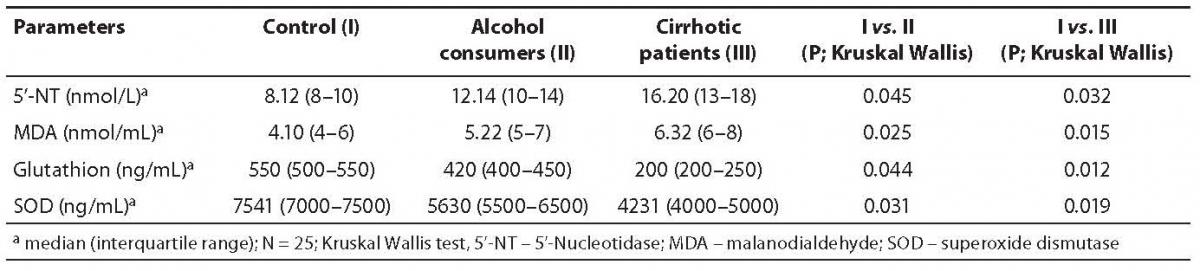
The RBC SOD values were also highest in the control group, relatively lower in alcohol consumer group and lowest in cirrhotic patient group, as shown in Table 1. There was statistically significant difference in glutathione concentration between three groups (P < 0.05).
Discussion
In this study, serum 5’-NT level, the product of lipid peroxidation and antioxidant level were compared among cirrhotic patients, alcohol consumers and normal age matched individuals as controls. The high level of 5’-NT in the cirrhotic group was suggestive of intrahepatic obstruction of bile canaliculi due to fibrosis as the result of liver cell injury. Its relatively higher activity in the alcohol consumer group found support in a previous report stating that higher nucleic acid catabolism in chronic alcohol toxicity leading to its elevation as the enzyme is related to the breakdown of nucleic acid, especially mRNA. Chronic alcoholism produces a wide spectrum of liver and other organ diseases depending on the amount and duration of alcohol intake. Hepatic effects range from fatty lesion to hepatitis and cirrhosis, and the activity of 5’-NT will also be ranging as per the damage done to the liver cells and bile canalicular membranes, resulting in mild to moderate biliary stasis. The 5’-NT enzyme is induced in alcoholism.
Of importance is the fact that higher 5’-NT activity in the serum of patients identified with a high probability for the presence of liver disease confirms that any increase in 5’-NT activity is highly specific for hepatobiliary disease (1,2). Higher levels of MDA in both cirrhotic patient and alcohol consumer groups were suggestive of higher oxidative stress in patients with liver cirrhosis as well as in alcohol consumers (23). These findings are consistent with the reports from several studies that confirmed the involvement of free radicals in the pathogenesis of liver injury in case of liver cirrhosis and chronic alcoholism (29-31). Glutathione is of major importance in the reduction of hydrogen peroxide and organic peroxides (e.g., lipid peroxides) in a reaction that is catalyzed by selenium containing GSH peroxidase and by other proteins that also exhibit GSH-S-transferase activity (32). It is the major endogenous antioxidant produced by the cell. It participates directly in the neutralization of free radicals, reactive oxygen compounds and maintains exogenous antioxidants such as vitamins C and E, and also plays a role in detoxification of many xenobiotics (23).
SOD plays a role in the removal of hydrogen peroxide (H2O2) formed in red cells and because hemoglobin and SOD have been shown to be in close association in red cells.
The low levels of GSH and SOD in cellular and extracellular fluids reduce their oxygen derived free radical scavenging capacity making the tissues more vulnerable to oxygen derived free radical (ODFR) damage (33). The low level of GSH and SOD in the cirrhotic patient and alcohol consumer groups was indicative of higher oxidative stress in patients with liver cirrhosis as well as in alcohol consumers (30).
The higher lipid peroxide levels (MDA) and lower antioxidant levels (GSH and SOD) in RBC of cirrhotic patients showed an altered oxidant and antioxidant status (30). Depletion of GSH renders the cell more susceptible to oxidative stress.
Our results demonstrated the antioxidant barrier to be impaired in liver cirrhosis, and to be associated with a decrease of glutathione level and the activity of the antioxidant enzyme SOD (18,19). Alcohol has been shown to deplete GSH levels, particularly in the mitochondria, which normally are characterized by high levels of GSH needed to eliminate the ROS generated during the respiratory chain activity.
Reduced glutathione in cirrhotic patient and alcohol consumer groups was consistent with other reports (19). This observation may be explained on the basis of (i) its utilization in scavenging free radicals, (ii) its involvement in maintaining non-GSH critical protein sulfhydryls in reduced state, (iii) acting as a co-factor for glutathione-S-transferase (GST) during detoxification of xenobiotics including alcohol, (iv) oxidation of glutathione to its oxidized form by glutathione peroxidase in detoxification of hydrogen peroxide and/or lipid peroxides, and (v) suppression of glutathione synthesis by ethanol. The more profound GSH reduction in alcohol cirrhotic patients may be due to the synergistic action of alcohol and liver cirrhosis.
The lowest level of glutathione found in the cirrhotic patient group indicated imbalance of the glutathione antioxidant system in cirrhosis, supporting the hypothesis that oxidative stress plays an important role in the development of liver cirrhosis (19). The low levels of SOD activity observed in the cirrhotic patient and alcohol consumergroups were indicative of oxidative stress that may be responsible for maximal destruction in liver architecture. A markedly lower SOD activity observed in the cirrhotic patient group indicated that oxidative stress was much more profound in liver cirrhosis patients than in alcohol consumers, as liver cirrhosis is a common outcome of a variety of chronic liver diseases (22).
A significant increase in MDA levels in the cirrhotic patient and alcohol consumer groups suggested that cirrhotic patients and alcohol consumers were at higher exposure to oxidative stress. The decreased levels of both GSH and SOD in the present study suggested that the increase in oxidative stress was associated with a proportionate decrease in the antioxidant defense system in cirrhotic patients and alcohol consumers.
From these findings, we can conclude that the activity of 5’-NT rises consistently in cirrhotic patients and alcohol consumers according to the extent of damage to the liver and that there is an increase in oxidative stress and a decrease of antioxidant status in both liver cirrhosis patients and alcohol consumers; however, the extent of oxidative stress is more profound in cirrhotic patients than in alcohol consumers.
Conclusion
This study attempted to establish the extent of serum 5’-NT activity in cirrhotic patients and alcohol consumers as compared with normal individuals (controls). This study also attempted to establish the extent of oxidative stress and antioxidant status in cirrhotic patients and alcohol consumers.
Based on study results, it is concluded that the activity of serum 5’-NT is consistently higher in cirrhotic patients and alcohol consumers, according to the extent of liver damage, hepatobiliary damage, and biliary stasis. We can also conclude that there is an increase in oxidative stress and a decrease in antioxidant status in both liver cirrhosis patients and alcohol consumers, whereby the extent of oxidative stress is more profound in cirrhotic patients than in alcohol consumers with or without a history of hepatobiliary disease. The parameters of lipid peroxidation and antioxidant defense may be useful markers for monitoring patients with hepatobiliary disorders.
Acknowledgment
The authors are grateful to Kasturba Medical College, Mangalore, for providing facilities to carry out these studies.